
CYKASY

Rod Encephalartos Lehm. v jižní Africe
R. A. Dyer, Botanical Research Institute, Pretoria
Živa 1975, 5: 168-169 a 6: 206-208 - připraveno ve spolupráci s časopisem Živa
Cykasy jsou pozoruhodná, velmi starobylá skupina rostlin. Vnějším vzhledem se podobají palmám, příbuzné však s nimi nejsou. Palmy, stejně jako ostatní krytosemenné rostliny, mají vajíčka uzavřena v plodolistech (semenících), zatímco u cykasů jsou vajíčka nekrytá a na plodolisty přisedají volně. Řadíme je proto mezi rostliny nahosemenné.*) Recentní cykasové rostliny bývají zpravidla členěny do tří čeledí. Liší se žilnatinou listů a uspořádáním samičích plodolistů:
-
a.lístky lichozpeřených listů pouze s jedinou střední silnou žilkou, postranní žilky chybějí; samičí plodolisty jednotlivé, netvoří šištice (Cycadaceae Lindl.)
-
b.lístky lichozpeřených listů s několika (až mnoha) podélnými, rovnoběžnými, víceméně stejně silnými žilkami; samičí plodolisty jsou nahloučeny v šišticích (Zamiaceae Reichenb.)
-
c.lístky lichozpeřených listů se zpeřenou žilnatinou, tj. se silnější střední žilkou a mnoha příčnými postranními žilkami: samičí plodolisty jsou v šišticích (Stangeriaceae Johnson.)
Pozn. autora webu: od doby publikace tohoto článku v roce 1975 bylo popsáno mnoho nových druhů, počty druhů v tomto článku uváděných již tedy neplatí. Přesné počty najdete v Atlase:
Čeleď Cycadaceae obsahuje jediný rod Cycas L. Více než 20 druhů se vyskytuje převážně v jihovýchodní Asii (na západ až po Indii, na jih po severovýchodní Austrálii, na sever po jižní Japonsko). Jediný druh zasahuje na Madagaskar a pobřeží východní Afriky.
Čeleď Zamiaceae zahrnuje osm rodů: Zamia L. s více než 15 druhy), Dioon Lindl. (s více než 15 druhy], Microcycas Miq. (1 druh), Ceratozamia Brongn. (25 druhů), Bowenia Hook. (2 druhy), Macrozamia Miq. (14 druhů), Lepidozamia Reg. (2 druhy) a Encephalartos Lehm. (více než 45 druhů). Rozšíření čeledi je mnohem větší než předešlé. První 4 rody jsou zastoupeny na obou amerických subkontinentech, následující tři jsou domácí v Austrálií. Druhým nejbohatším rodem je však Encephalartos, vyskytující se v jižní a tropické Africe. Pozn autora webu: v současnosti je do této čeledi řazen i rod Chigua se dvěma druhy.
Čeleď Stangeriaceae tvoří jediný rod Stangeria T. Moore s monotypickým druhem S. eriopus (Kunze) Nash, domácím v jižní Africe. Pozn autora webu: v současnosti je do této čeledi řazen i rod Bowenia se dvěma druhy.
Dnešní cykasové rostliny obsahují tedy něco málo přes 120 druhů (pozn. autora webu: v současnosti již více než 300) v 10 rodech a představují malou přežívající skupinu archaických rostlin. Na Zemi se objevily nejméně před 250 milióny lety. V období před 200-100 milióny let byly jednou z hlavních složek tehdejší vegetace. Posledních 50 miliónů let přežily prakticky bez jakýchkoli podstatnějších změn. Proto se o nich mnohdy hovoří jako o žijících fosiliích.
Pohromu pro cykasy přinesl zejména rozvoj krytosemenných rostlin. Soupeření o životní prostor na Zemí se postupně neustále stupňovalo a po vzniku travnatých společenstev přistoupil navíc další činitel, nejničivější ze všech přírodních živlů, oheň. Konkurence krytosemenných rostlin drasticky zmenšila počet cykasů i jejich areály, zatlačila je na jejich dnešní, nepříliš nápadné místo mezi současným rostlinstvem, jejich úplný zánik však nepřinesla. Přežily především druhy, které mají účinná ochranná zařízení proti požárům: je to buď podzemní, málo vyvinutý stonek, nebo nadzemní kmen, chráněný speciální nehořlavou vrstvou.
Ochranná vrstva vzniká z bází řapíků (pokrytých chlupy a šupinkami) postupně opadávajících starých listů. Na kmenu se střídají velmi těsně nahloučené báze odumřelých listů s oválnými nebo kopinatými šupinami a tvoří nehořlavou, zhruba 2—8 cm tlustou izolační vrstvu, která spolehlivě chrání živé tkáně stonku před požáry běžné intenzity. Jakékoli mechanické poškození ochranné vrstvy však znamená vážné ohrožení života celé rostliny.
Další okolností, která mohla anebo nemusela mít vliv na přežití cykasů do dnešních dnů, je jedovatost většiny druhů. Jak vegetativní orgány (listy, kmeny i kořeny], tak i semena bývají často silně toxická.
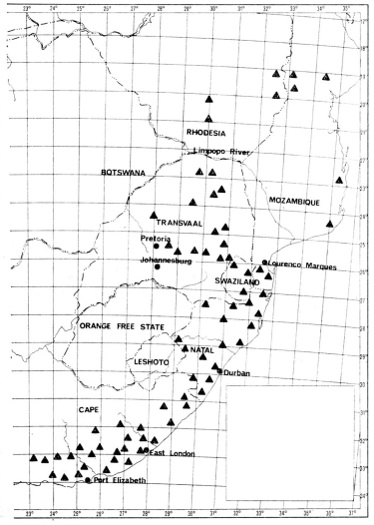
V Africe jsou cykasové rostliny zastoupeny — s výjimkou pouhých dvou druhů z rodů Cycas a Stangeria — jen druhy poměrně bohatého rodu Encephalartos. Jejich rozšíření na jihu kontinentu podává naše mapka.
Cykasy rodu Encephalartos jsou vždy dvojdomé (exempláře jsou odděleného pohlaví). Kořen je v mládí hlízovitý, později se vytvářejí i postranní jemnější, větvící se kořeny.**)
Stonek (kmen) je buď silně zakrnělý, podzemní nebo slabě nad povrch půdy vyčnívající, anebo u jiných druhů vzpřímený, až 10 (-15) m vysoký. Na bázi rostliny se mohou často tvořit postranní odnože, nadzemní kmen se však větví jen vzácně (u starých jedinců nebo po poškození vrcholového meristému). Kmen je vždy pokryt spirálovitě uspořádanými šupinami a bázemi odumřelých listů. Asimilační listy, 40 až 250 cm dlouhé, jsou nahloučeny ve vrcholovém chocholu. Může jich být malý počet, ale někdy i několik desítek. Jsou produkovány střídavě s generativními orgány, většinou nepravidelně (i v několikaročních intervalech) a v závislosti na růstových podmínkách. Listy mají silné pružné střední žebro a velký počet tuhých, kožovitých lístků.
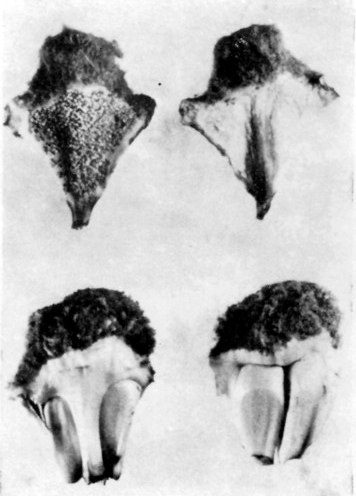
Plodolisty (sporofyly) jsou v šišticích. Samčí šištice nesou na spodní straně plo-dolistů prašná pouzdra [mikrosporangia] s pylovými zrny. Plodolisty samičích šištic mají na svrchní straně dvě velká vajíčka, proti nepříznivým vnějším vlivům chráněná přerostlou štítkovitou částí plodolistu. Vejčitá až elipsoidní semena jsou obalena více nebo méně masitým vnějším osemením [sarkotestou] a tvrdým vnitřním osemením [sklerotestou], chránícím nepatrné embryo, uložené uvnitř bohatého endospermu.
Rod Encephalartos má podle současných znalostí asi 45 druhů. Z nich 28 roste v jižní Africe jižně od řeky Limpopo, ostatní jsou rozptýleny na území tropické Afriky (severní hranicí rozšíření rodu je Sahara). Velká část těchto druhů se dnes zachovala už jen na omezeném území a během několika desetiletí, po něž tyto rostliny sleduji, jejich množství na jednotlivých lokalitách zřetelně pokleslo. Abychom dokázali jejich dnešní populace zachovat a pokud možno i zvětšit, snažíme se vysvětlit řadu otázek souvisejících s jejich ontogenezí.
Otázkou prvořadého významu je objasnění všech podmínek a požadavků úspěšné reprodukce v přírodě. Jak u těchto rostlin nastává opylení? Na první pohled jde o triviální záležitost, přesto však nebyly doposud všechny detaily dostatečně podrobně vysvětleny. Dosud existují rozdílné názory i na způsob opylení: jedni tvrdí, že cykasy jsou větrosprašné, jiní dokazují, že jsou hmyzosprašné. Opylení může být jistě zprostředkováno oběma způsoby, ale domnívám se, že opylení větrem je častější. Pozorovali jsme, že v přírodě jsou vždy šištice buď celé plodné, anebo celé úplně sterilní. Kdyby se na opylení podílel hmyz, stěží by k tomu mohlo dojít. Zdá se, že nevane-li ve vhodném období vítr určitým směrem a určitou rychlostí, pak k opylení nedojde. K pozitivnímu výsledku je však ještě zapotřebí, aby samčí a samičí exempláře rostly v určitém vzájemném poměru a navíc i v určitém množství a hustotě. Poklesne-li hustota populace pod určitou kritickou hodnotu, nemůže být už opylení zaručeno a je ohrožena přirozená reprodukce a tím samozřejmě i samotná existence této populace. (Pozn autora webu: v současnosti jsou již cykasy považovány za většinově opylované hmyzem, byť s výjimkami především u rodu Cycas)
Při pokusech s umělým opylením (jak normálním pylem, tak emulzí pylu ve vodě} dosáhli jsme dobrých výsledků: semena byla nejen dobře vyvinuta, ale i klíčivá. V přírodě se totiž semena tvoří většinou i v případě, že vajíčka opylena a oplozena nebyla. Embryo (se spirálovitě utvářeným suspenzorem) v neoplozených semenech pochopitelně chybí, vyvíjí se pouze škrobnatý endosperm s testou, semena tedy nemohou klíčit a dát vznik novému jedinci.
Vytváření šištic není proces jednotný ani v rámci rodu, dokonce ani u druhu nebo u téhož jedince. Samčí strobily se vyvíjejí poměrně rychle, zpočátku jako terminální orgány, a do 1,5—3 měsíců vytvářejí pylová zrna. Po uvolnění pylu z prašných pouzder rychle zasychají a opadávají. Naproti tomu samičí strobily zůstávají po opylení další 3—9 měsíců bez viditelné změny. Semena postupně zrají, bez ohledu na to, jsou-li oplozena (tj. klíčivá), anebo neoplozená (tj. neklíčivá). Potom se postupně samičí strobily rozpadávají (viz snímek) a uvolňují semena, která dopadají do bezprostřední blízkosti mateřské rostliny. Semena dosahují u některých druhů velikosti i přes 3 cm (jsou však mnohem menší než u rodu Cycas, kde dosahují až přes 8 cm délky). Klidové stadium neexistuje, embryo pokračuje v pomalém růstu uvnitř endospermu. Toto živné pletivo zaručuje při své velikosti dostatečnou zásobu živin i vody, ovšem semeno přesto nesmí po dozrání nikdy úplně vyschnout. Klíčí jen ta semena, která zapadla do substrátu, udržujícího mírnou vlhkost.
Dalších asi 6—12 měsíců nejsou na semenech patrné viditelné změny. Teprve pak se objevuje první část nové rostliny - hlízovitý primární kořen. Vyvíjí se velice pomalu, opět v závislosti na podmínkách prostředí. Rychlost je různá i u jednotlivých druhů. Celkem 7—15 let trvá, než vyvíjející se stonek dosáhne velikosti tenisového míčku. Dalších 30 až 50 let musí uběhnout, než dosáhne velikostí fotbalového míče. Potom, když už je dosaženo přibližně průměru kmene dospělého jedince, dochází k prodlužování stonku. V té době je také dosaženo optimálního počtu listů a další přírůstky se stávají pravidelnějšími (odhlédne-me-li od klimatických výkyvů). Prodlužování kmene se ovšem týká jen druhů s vyššími nadzemními kmeny. Ty také mívají poměrně velké roční přírůstky, tj. 1—3 cm výšky za rok. Druhy s podzemním stonkem se s těmito obřími příbuznými měřit nemohou; jejich roční přírůstky činí jen zlomky milimetrů.
Bohužel existuje jen málo záznamů, z nichž by bylo možno vypočítat nebo odhadnout skutečnou rychlost růstu cykasů. Výjimkou jsou snímky cykasů Encephalartos altensteinii Lehm., získané v roce 1915, které pomohly utvořit představu o celkovém přírůstku za 6 desetiletí (viz snímek). Porovnáním jsme dospěli k výsledku, že 105 cm celkového přírůstku představuje téměř 1,5 cm za rok. Musíme ovšem počítat i s tím, že v kultuře (jde o exempláře v botanické zahradě v Durbanu) jsou podmínky příznivější než v přírodě (hlavně pokud jde o dostatek vody), a tak by zjištěný závěr mohl popř. vést i k chybnému zobecnění.
Impozantní exemplář cykasu Encephalartos eugene-maraisii Verdoorn, jehož stáří, počítáme-li je od vyklíčení semena, je nutno odhadovat na tisíciletí, zachycuje jeden z uveřejněných snímků. Větší větvící se kmen vlevo, jevící jíž známky senility, může být přibližně 600 let starý, velký vzpřímený kmen napravo přibližně 400 - 500 let. Během času tyto kmeny odumřou, ale v té době již budou nahrazeny bazálnímí odnožemi, I nichž největší postupně nahradí starý kmen a dosáhne podobných rozměrů. Můžeme tedy usuzovat, že spodina rostliny je mnohem starší než vyšší nadzemní kmen, a není-li tento předpoklad chybný, pak odhad tisíciletého stáří báze individua není přehnaný.
Naproti tomu E. transvenosus Stapf et Davy netvoří v přírodě odnože, nemůže se zmlazovat a regenerace jeho populace plně závisí na produkci klíčivých semen. Je to nejvyšší jihoafrický druh. Stáří desetimetrového jedince na našem snímku se odhaduje na 500 let. Tento druh ze severovýchodního Transvaalu (území Eantu) osidluje pobřežní pás země, bohatý na mlhy, s vlhkým přímořským vzduchem. Královna Modjadji je často titulována „královna dešt”, protože údajně dokáže vládnout živlům a je schopna přivolat déšť v období, kdy jej plodiny nejvíce potřebují. Není sice známo, z jakých důvodů je k tomuto „ovlivňování počasí" zapotřebí cykasů, ale faktem zůstává, že její královská autorita (stejně jako předchozích členů dynastie) postačuje k tomu, aby domorodci ochraňovali a ošetřovali jedinečný les nádherných exemplářů tohoto druhu.
Je-li E. transvenosus na svém nalezišti poměrně hojný, pak E. woodii Sander je skutečně jedinečně vzácný v plném smyslu tohoto slova. Roku 1895 ho nalezl M. Wood v Zulsku v jediném exempláři. Samčí rostlinu tvoří čtyři silné kmeny, asi 5,5 m vysoké. Jde doposud o jediný nález tohoto druhu v přírodě. Již M. Wood uvedl, že v blízkosti nerostl jiný exemplář a ani později nebyla podobná rostlina bohužel již nikdy nalezena. Bohužel! Neboť je to snad nejkrásnější ze všech druhů rodu, má nejozdobnější, deštníkovitý chochol listů. Počátkem tohoto století bylo od mateřské rostliny odděleno několik mladých postranních odnoží a v Anglii byl podle nich druh popsán a zaveden do kultury (viz snímek). V současnosti jsme vynaložili velkou námahu ve snaze nalézt druhý exemplář samičího pohlaví pro možnost generativního rozmnožení druhu, ale všechny snahy byly opět zcela bezúspěšné. Nelze ani zjistit, zda jde o mutanta, vzniklého z poměrně vzdáleně podobného E. natalensis, nebo o relikt prehistorických dob.
Další ze snímku zachycuje původní porost E. natalensis Dyer et Verdoorn na útesech a skalních stěnách údolí v Natalu. Druh nebyl nikdy nalezen výše než v 700 m n. m. Ekologický charakter ukazuje na jeho blízkou příbuznost s E. altensteinii Lehm.; tento druh sestupuje až na úroveň mořské hladiny v říčních údolích východního Kapská.
E. ghellinckii Lehm. se nápadně liší od všech ostatních druhů úzkými převislými lístky (viz snímek). Je porůznu nalézán v pohoří Drakensberg v Natalu, kde vystupuje do vyšších poloh než kterýkoli jiný druh rodu, tj. do 2000 m n. m. Při horní hranici svého výskytu je každoročně vystaven sněhovým srážkám a nízkým teplotám, které snáší bez poškození. Ohnuté, převislé postavení lístků usnadňuje opadávání a odtávání sněhu, pokud ho není příliš mnoho; potom sníh pokrývá celé rostliny. Kromě vysoké tolerance k nízkým teplotám se u E. ghellinckii setkáváme s mnohem silnější izolační vrstvou odumřelých bází řapíků a šupin kolem kmene. Ta slouží nejen k izolaci tepelné, ale i k ochraně proti požárům, které jsou v horských porostech poměrně Časté zčernání a zuhelnatění povrchu kmene, ostře kontrastující s barvou sněhu, ukazuje uveřejněný snímek. Téměř zralé šištice E. ghellinckii jsou silně vlnaté a jsou podobné šišticím některých jiných druhů, např. E. lanatus. U E. lanatus Stapf et Davy pozorujeme nápadně rozdílné utváření šištic obou pohlaví (viz snímky samčích a samičích strobilů); jsou na první pohled rozlišitelné. Samčí strobily bývají také na rostlinách méně početné než samičí.
E. heenanii Dyer je naproti tomu zástupcem skupiny druhů, kde vnější vzhled šištic je u obou pohlaví prakticky stejný. Obojí šištice mají šupiny stejných rozměrů, stejného uspořádání i stejného zbarvení povrchu. Pohlaví šištic však okamžitě zjistíme, vezmeme-li je do ruky. Samičí, hustě napěchované velkými semeny, jsou vždy zřetelně těžší než samčí, s poměrně velkými vzduchovými prostorami mezi šupinami.
Na pohled nejpůsobivější africké cykasy jsou nepochybně právě nejvyšší druhy rodu Encephalartos. Jejich mohutné, vysoké kmeny nesou obrovské chocholy dlouhých listů. Některé z nich, jako E. altensteinii, E. natalensis, E. woodii a E. transvenosus, byly již zmíněny. Nelze však opomenout E. longifolius (Jacq.) Lehm. (viz snímek), první známý druh rodu, který už v roce 1773 v Jižní Africe nalezl známý švédský botanik Karl Peter Thunberg. Zamýšlel ho pojmenovat E. caffer, ale pak došlo k záměně: v okolí výskytu E. longifolius (východní Kapsko) roste i jiný, miniaturní druh, který dnes nese původní označení E. caffer (Tlumit.) Dyer. Thunberg popsal šištice malého druhu a omylem je pokládal za strobily vysokého stromovitého druhu (dnešní E. longifolius). Omyl nebyl dlouho rozpoznán a teprve nedávno byl opraven. Byl to však právě Thunberg, který jako první zaznamenal, že domorodci používají škrobem bohaté dřeně kmene k přípravě primitivního chleba. Z toho povstal i název této skupiny cykasů „Kaffirbread trees**, podobně jako řecký ekvivalent Encephalartos („chléb v hlavě"). Metoda, používaná původním obyvatelstvem, spočívala v otevření kmene a vybrání střední dřeně, která pak byla zabalena do živočišné kůže a na dobu asi 6 týdnů zahrabána do jámy. V půdě nastala částečná fermentace a rozrušení husté vláknité struktury dřeně, jež potom mohla být již snadno rozdrcena mezi kameny a s vodou uhnětena v těsto. Příprava chleba, jak poznamenává Thunberg, velmi bídného chleba, byla dovršena pečením a pražením.
Přestože, jak bylo v úvodu řečeno, Cykasy se během posledních 50 miliónů let zásadně nezměnily, některé současné druhy mají přece jen určité modifikace, dané sice pomalým, ale přesto existujícím vývojem. U E. horridus (Jacq.) Lehm. pozorujeme přizpůsobení stále suššímu a suššímu klimatu jeho naleziště. Roste v blízkosti Uitenhage ve východním Kapsku ve formaci nízkého křovinatého buše, který má vysloveně xerofytní charakter. Na lokalitě je zastoupeno mnoho sukulentů s podzemními cibulemi nebo hlízami a řada sukulentů listových a stonkových. E. horridus nese znaky přizpůsobení ekologickým podmínkám: poměrně malé, ale tuhé lístky, hluboce vykrajované, ostře zubaté, s modrozeleným lesklým povrchem umožňujícím odraz části dopadajícího slunečního záření. Náš snímek však zachycuje rostlinu pěstovanou v botanické zahradě, která zřejmě ve výhodnějších životních podmínkách, než má v přírodě, podstatně změnila utváření kmene (roste rychle a tvoří množství postranních odnoží).
Lokality některých druhů leží poměrně blízko sebe a známe dokonce případy kdy se dva druhy vyskytují na jednom nalezišti společně; např. E. longifolius a E. horridus u Uitenhage. To dovoluje i předpoklad možnosti vzájemné hybridizace. Na zmíněné lokalitě byly skutečně nalezeny exempláře, které jsou zjevnými kříženci mezi oběma uvedenými druhy. Podobně i E. altensteinii má příležitost i schopnost křížit se s E. trispinosus (Hook.) Dyer, E. latifrons Lehm. a E. villosus Lem. (což jsou poměrně značně odlišné samostatné druhy).
Není mým zvykem pózovat při fotografování, ale abych dodal tomuto příspěvku trochu osobního, přátelského pozadí, zařadil jsem i svůj snímek s E. eugene-maraisii v přirozeném porostu v horách Waterberg v severním Transvaalu. Je třeba jen dodat, že od doby, kdy byl exponován, uběhlo 30 let.
Poznámky
*) Nahosemenné rostliny nelze pokládat za příbuzensky jednotnou skupinu, nýbrž pouze za určitý vývojový stupeň různorodých evolučních větví; cykasové rostliny jsou pokládány za naprosto samostatnou vývojovou linii (zcela nepříbuznou jehličnanům, chvojníkům i rostlinám ginkgovitým), která se už od konce paleofytika vyvíjela nezávisle a maxima rozvoje dosáhla v juře a ve spodní křídě. Pozn. red. J. S.
**) U některých druhů rodu Encephalartos byly na kořenech pozorovány malé nádorky, ve kterých, podobně jako u jiných rostlin (např. u čeledi Viciaceae) žijí symbioticky baktérie schopné asimilovat atmosférický dusík a přeměňovat jej v biologicky využitelnější formy, např. na dusičnany. Pozn. překl.
Obrázek
Klíčení semen rodu Encephalartos: a, b, c — podélný řez semenem v různých stadiích vývoje; d, e — stadia, kdy se objevuje první kořen a první list. Semena bez spirálovitého útvaru (a, b) jsou sterilní (bez zárodku — neoplozená)








Fotografie
Dvouleté semenáče rodu Encephalartos ve školce Ochrany přírody v Pretorii
Obrázek
Rozšíření rodu Encephalartos v jižní Africe
Fotografie
Encephalatos ghellinckii
Fotografie
Encephalartos horridus. Jedinec pěstovaný v botanické zahradě, kam byl přenesen z přírodních podmínek v okolí Vredehuisu.
Fotografie
Samčí a samičí plodolisty - nahoře samčí, dole samičí, vždy vlevo abaxiální a vpravo adaxiální strana
Fotografie
Samičí rostlina Encephalartos lanatus
Fotografie
Encephalartos friderici-guilielmi - velmi starý exemplář ve východním Kapsku s ohořelými zbytky řapíků
Fotografie
Encephalartos ghellinckii pod sněhovou pokrývkou v pohoří Drakensberg
Fotografie
Desetimetrový jedinec Encephalartos longifolius poblíž Driwelskloofu
Fotografie
Encephalartos longifolius
Všechny snímky R. A. Dyer